
Какие изменения могут быть полезными для эволюции

Следует выделить четыре основных фактора эволюции: мутационный процесс, изоляция, популяционные волны (и дрейф генов),
а также единственный направленный фактор эволюции — естественный отбор.
Мутационный процесс
Мутации (лат. mutatio — изменение) — стойкое изменение генома (наследственного материала), которое может быть унаследовано
потомками организма. Процесс возникновения мутаций — мутагенез.
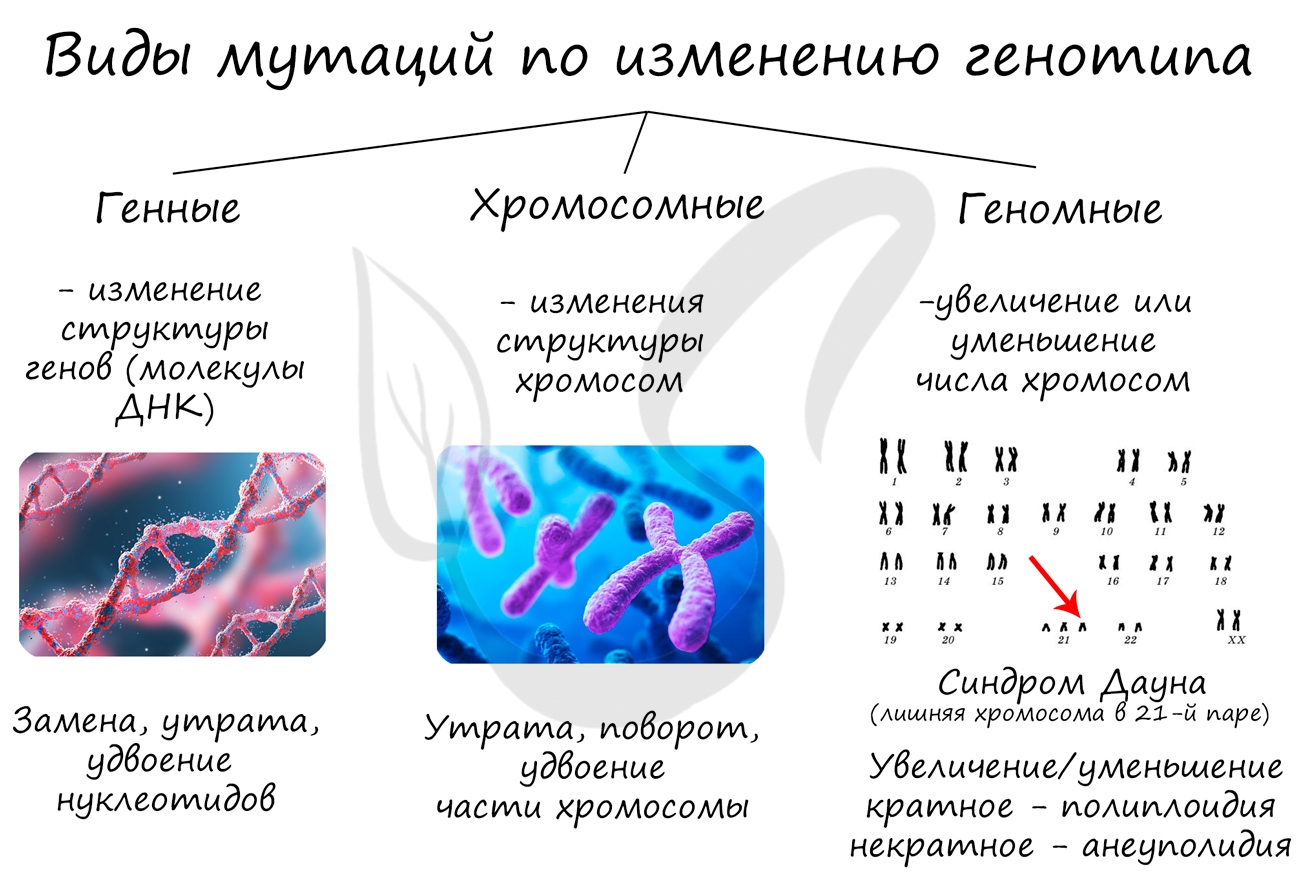
Мутации подразделяются на:
- Генные
- Хромосомные
- Геномные
Большинство мутаций возникает спонтанно и вредит организму. Часть мутаций являются рецессивными, поэтому не проявляются
и передаются многим поколениям, накапливаясь в генофонде популяции.
Мутации напоминают колоду карт: неизвестно, что выпадет — чаще всего это карты невысокого ранга, козыри — большая удача. Так
и мутации, большинство из них вредные, приводят к развитию опухолей. Полезные встречаются гораздо реже. Как игра в карты,
все подчиняется случайности.

Популяционные волны
Все виды подвергаются воздействию периодически возникающих популяционных волн — уменьшению или увеличению численности особей.
Популяционные волны могут возникать по ряду причин:
- Сезонные изменения, периодические изменения какого-либо значимого фактора среды
- Непериодические изменения, например, в результате природных катастроф, изменение численности популяций хищник-жертва
- Заселение новых территорий, ярким примером которого является бурный рост численности кроликов, завезенных в Австралию
Колебание численности популяций по типу «хищник — жертва» является классическим примером популяционных волн. Представим себе
популяцию зайцев (жертв), которая бурно увеличилась в численности. Зайцами питаются лисица, волк (хищники). С увеличением
их кормовой базы (зайцев) наблюдается и рост численности хищников, которые поедают зайцев, вследствие чего численность зайцев
снижается. С уменьшением кормовой базы, снижается и число хищников. Так в природе устанавливается баланс между хищниками и
жертвами.

Особенно весомым фактором эволюции популяционные волны выступают в небольших популяциях. Их участие в эволюционном процессе
основано на явлении дрейфа генов.
Форма гена — аллель, с которым вы подробнее познакомитесь в ходе изучения генетики, встречается в популяции с определенной
частотой. Дрейф генов — изменение частоты встречаемости аллельных вариантов генов.
Дрейф генов обусловлен случайными причинами: у особей образуются гаметы, несущие различные формы аллельных генов. Не все
из гамет принимают участие в процессе оплодотворения: здесь вновь руководит случайность. Вследствие этого одни аллельные
формы генов могут встречаться в популяции часто, другие — редко.
Если представить, что часть особей, составляющих одну популяцию, погибли по тем или иным причинам, то редкие гены в оставшихся
особях могут увеличить свою частоту, то есть в результате размножения оставшихся особей редкие гены начнут встречаться более часто
— это и есть дрейф генов.
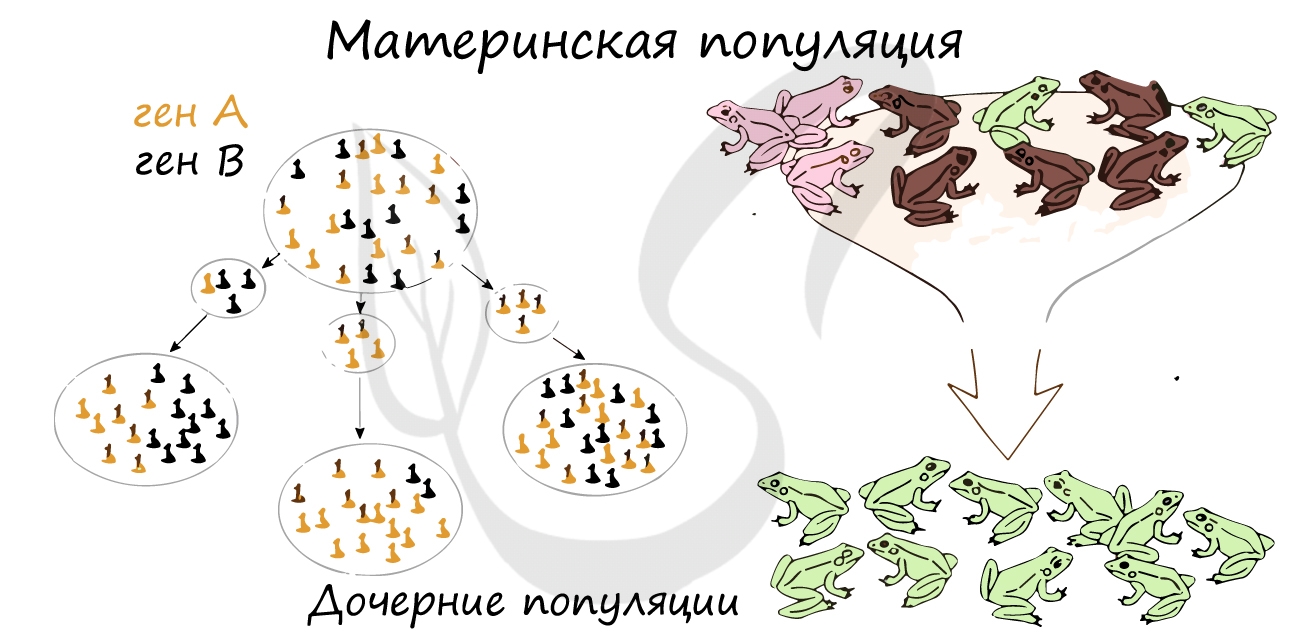
В закрытых популяциях не только животных, но и людей — в религиозных общинах, происходит возрастание гомозиготности популяции,
что приводит к снижению ее жизнеспособности и проявлению редких аллелей.
Такое повышение встречаемости аллелей возникает в результате
близкородственных браков: проявляются редкие гены, которые часто приводят к заболеваниям.

Изоляция
Изоляцией называют невозможность или затруднение свободного скрещивания между особями одного вида. Вследствие этого, генофонды
двух популяций становятся независимыми друг от друга. Внутри каждой популяции происходит генотипическая дифференцировка из-за
их разобщенности.
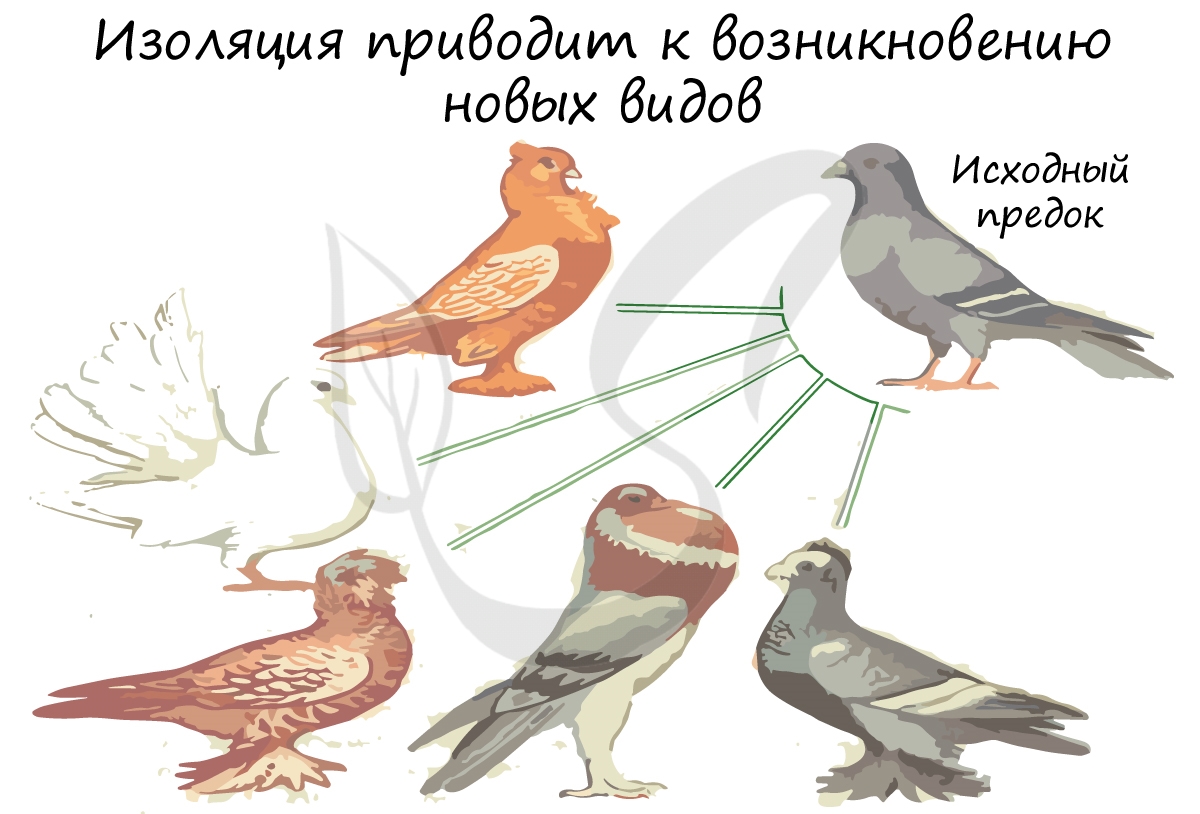
Популяции приобретают все больше различий, и в результате действия естественного отбора из этих популяций могут образоваться
различные новые виды. Выделяется два вида изоляции:
- Географическая (пространственная)
- Биологическая (экологическая, временная)
- Экологическая — особи обитают на одной территории, но в различных местах обитания (к примеру, разделены друг от
друга непроходимой чащей) - Временная — изоляция вследствие разновременности половой активности, периода цветения
- Этологическая — изоляция вследствие различного брачного поведения
- Механическая — отличия в строении половых органов, невозможность спаривания
Географическая (греч. geo — земля) изоляция может возникать вследствие географических барьеров — пустыни, горы, водоемы.

Естественный отбор
Изученные нами факторы эволюции: мутации, популяционные волны и дрейф генов, изоляция — все они носят случайный, ненаправленный
характер. Они приводят к появлению различных признаков у отдельных особей, которые могут быть как полезны, нейтральны, так и вредны для
особи.
Таким образом, перечисленные факторы создают основу, «базу» для действия единственного направленного фактора эволюции — естественного
отбора. В ходе естественного отбора особи с полезными признаками, которые помогают им приспособиться к условиям внешней среды и
способствуют выживанию, остаются и размножаются, а особи без этих признаков выживают реже и не продолжают род.

Закон естественного отбора безапелляционно провозглашает: будь приспособлен — или умри. Выживает в природе не самый сильный,
а самый приспособленный. Иногда выжить животным помогает и сила, но гораздо больше других примеров. Многие животные сливаются
с окружающей средой: приобретают покровительственную окраску (мимикрию), которая делает их незаметными.
Иногда безобидные животные, в результате приспособления к внешней среде, приобретают окраску тела, напоминающую окраску опасных
хищных животных. Примером может послужить внешнее сходство мухи из семейства журчалок с осой.

Многие хорошо защищенные, ядовитые виды в ходе естественного отбора получили яркую, так называемую предупреждающую окраску.
Эта окраска предупреждает хищников об опасности. Если хищник съест такое ядовитое животное, то рискует получить тяжелую
интоксикацию и погибнуть.

Теперь вы понимаете, что признаки животных — различные формы их тела и окраска — являются приспособлениями к условиям внешней среды,
это — полезные признаки, которые в ходе естественного отбора позволили животным выжить и размножиться. Таким образом, естественный
отбор это отбор особей, с наиболее приспособленным к среде фенотипом.
Необходимо осознавать относительность приспособленности к окружающей среде. Она помогает выживать лишь при определенных условиях, и,
если условия меняются, то окраска может оказаться вовсе не полезной, но даже и вредной. К примеру, при таянии снега заяц-беляк становится
еще более заметен на голой земле.

Действие естественного отбора обусловлено несколькими факторами:
- Генетическое разнообразие особей, на основе которого возникают различные признаки
- Способность к неограниченному размножению (избыточность потомства)
- Борьба за существование
Если первые два фактора очевидны, то третий нуждается в более детальном рассмотрении. Чарльз Дарвин выделял несколько
форм борьбы за существование:
- Внутривидовая борьба
- Межвидовая борьба
- Борьба с неблагоприятными условиями окружающей среды
Самая ожесточенная борьба. Происходит между особями, принадлежащими к одному виду. Благодаря внутривидовой
борьбе происходит половой отбор: к размножению редко допускаются неприспособленные особи, род продолжают
лучшие из лучших.
Возникает между особями, которые принадлежат к разным видам. Более приспособленный к условиям среды вид побеждает и размножается, менее приспособленный — проигрывает и вымирает. Примером могут послужить формы взаимодействий: хозяин-паразит,
хищник-жертва, симбиоз.
В изменяющихся условиях внешней среды выживают наиболее приспособленные особи. Примером такой борьбы являются
сезонные миграции птиц, зимняя спячка у животных.

Формы естественного отбора
Различают несколько форм отбора:
- Стабилизирующий
- C узкой нормой реакции — цвет глаз, число пальцев у человека, окраска цветов растения
- C широкой нормой реакции — рост и вес человека, размеры листьев растения
- Движущий
- Дизруптивный (лат. disruptus — разорванный)
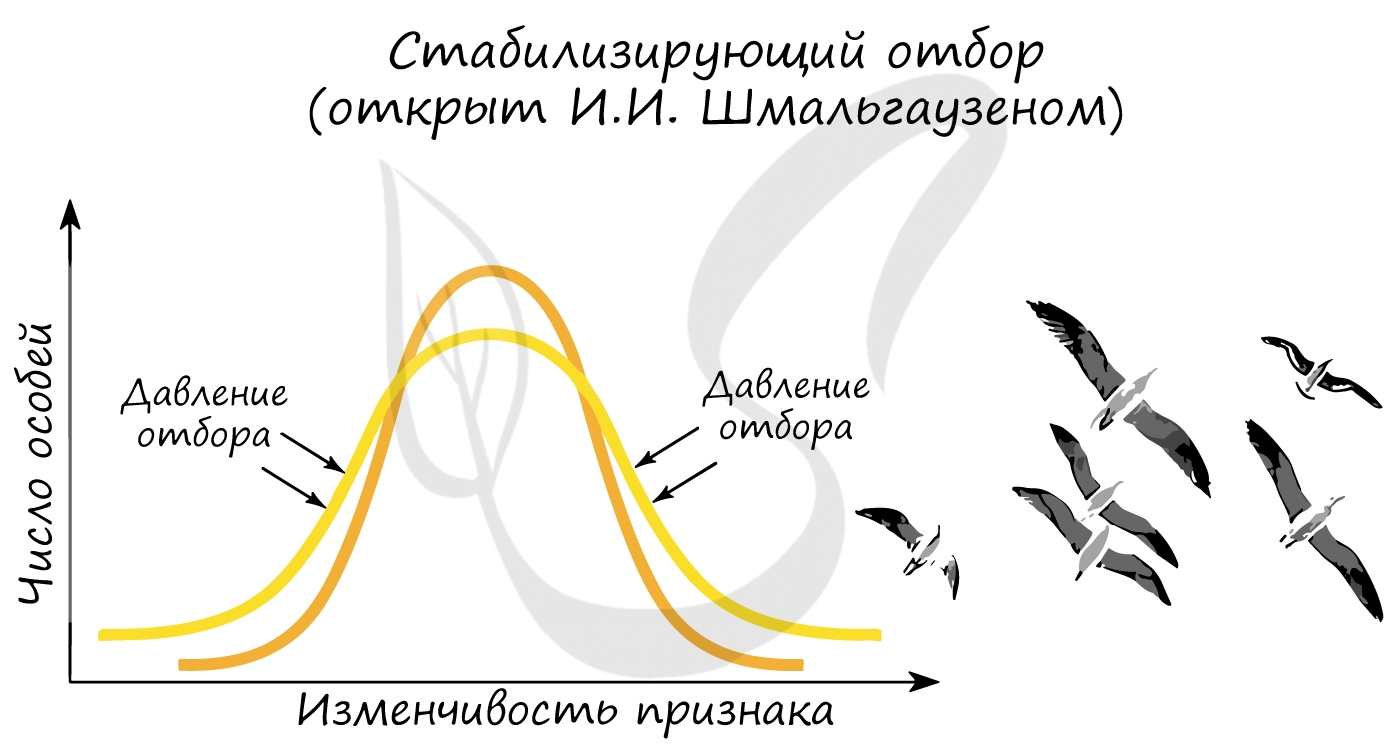
Открыт И.И. Шмальгаузеном. Стабилизирующий отбор приводит к сужению нормы реакции, устраняя отклонения от нее. В результате
преимущество получают особи, обладающие средней степенью признака, который характерен для вида или популяции. Этот отбор
действует при стабильных (неизменных) условиях среды.
Примером действия стабилизирующего отбора может послужить буря: во время бури чаще всего выживают птицы со средней длиной крыла, тогда как особи
с слишком короткими, или слишком длинными крыльями погибают.
Новый термин, который вы увидели — норма реакции — подразумевает способность генотипа, в зависимости от условий
среды, формировать различные фенотипы.
Чем шире норма реакции, тем более интенсивное влияние оказывает среда на признак. Признаки подразделяются на:
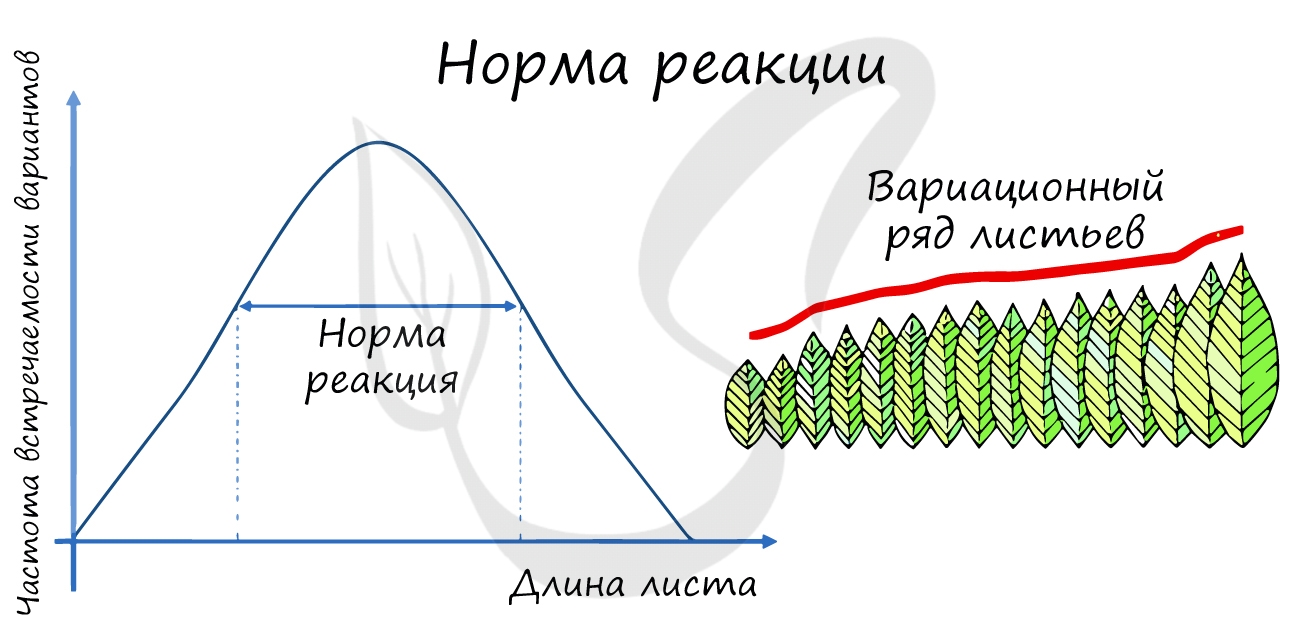
Движущий естественный отбор приводит к смещению нормы реакции, в результате чего изменяется среднее значение признака. Этот вид
отбора действует при изменяющихся условиях среды.
Известным примером является индустриальный меланизм — возникновение меланистических форм животных (греч. melanos — чёрный),
отличающихся темным окрасом. Это явление началось в Англии со второй половины XIX века вследствие бурного развития промышленности.
Из-за копоти, оседающей на поверхности стволов деревьев, бабочки со светлой окраской — берёзовые пяденицы — стали заметны
на стволах деревьев и легко поедались птицами. В результате остались только приспособленные — бабочки с темным окрасом,
которые были незаметны на стволах деревьев, вследствие чего они выживали и размножались.
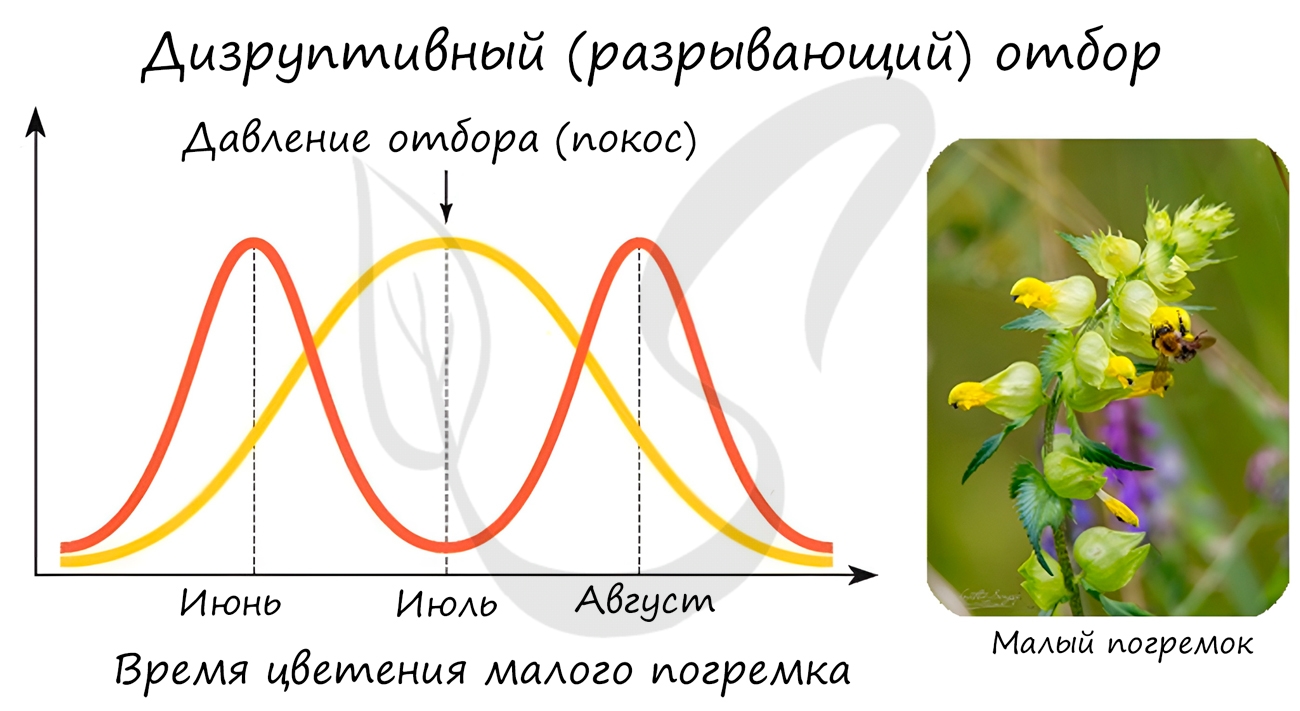
Направлен на сохранение в популяции крайних значений признаков, не благоприятствует среднему промежуточному значению
признака. В результате в популяции сохраняется более чем одно значение признака.
Типичным примером является появление в луговых сообществах раноцветущих и поздноцветущих растений. В результате летних покосов, особи
со средним значением признака, у которых цветение приходит на середину лета, постепенно исчезают из популяции растений. Выживают
и размножаются только те растения, у которых цветение происходит до или после покосов.
© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
В процессе исторического развития одни виды вымирают, другие изменяются и дают начало новым видам. Что же собой представляют виды? Существуют ли виды реально в природе?
Впервые термин «вид» ввел английский ботаник Джон Рей (1628- 1705). Шведский ботаник К. Линней рассматривал вид в качестве основной систематической единицы. Он не был сторонником эволюционных воззрений и считал, что виды со временем не изменяются.
Ж. Б. Ламарк отмечал, что различия между некоторыми видами очень незначительны, и в этом случае выделить виды довольно сложно. Он сделал вывод о том, что виды в природе не существуют, а систематика придумана человеком для удобства. Реально существует только особь. Органический мир представляет собой совокупность особей, связанных между собой родственны ми узами.
Как видно, взгляды Линнея и Ламарка на реальное существование вида были прямо противоположными: Линней считал, чтo виды существуют, они неизменны; Ламарк отрицал реальное существование видов в природе.
В настоящее время общепринята точка зрения Ч. Дарвина: виды реально существуют в природе, но постоянство их относительно; виды возникают, развиваются, а затем либо исчезают, либо изменяются, порождая новые виды.
Вид — это надорганизменная форма существования живой природы. Он представляет собой совокупность морфологически и физиологически сходных особей, свободно между собой скрещивающихся и дающих плодовитое потомство, занимающих определенный ареал и обитающих в сходных экологических условиях. Виды различаются по многим критериям. Критерии, по которым особи относятся к одному виду, представлены в таблице.
При определении принадлежности особи к какому-либо виду нельзя ограничиваться лишь одним критерием, а необходимо использовать всю совокупность критериев. Так, не возможно ограничиться только морфологическим критерием, поскольку особи одного вида могут различаться внешне. Например, у многих птиц — воробьев, снегирей, фазанов самцы внешне значительно отличаются от самок.
В природе у животных широко распространен альбинизм, при котором в клетках отдельных особей в результате мутации нарушается синтез пигмента. Животные с такими мутациями имеют белую окраску. Глаза у них красные, потому что в радужной оболочке нет пигмента, и сквозь нее просвечивают кровеносные сосуды. Несмотря на внешние отличия, такие особи, например белые вороны, мыши, ежи, тигры, относятся к своим видам, а не выделяются в самостоятельные виды.
В природе существуют внешне почти неразличимые виды-двойники. Так, раньше малярийным комаром называли фактически шесть видов, похожих внешне, но не скрещивающихся между собой и различающихся по другим критериям. Однако из них только один вид питается кровью человека и разносит малярию.
Процессы жизнедеятельности у разных видов часто протекают очень сходно. Это говорит об относительности физиологического критерия. Например, у некоторых видов арктических рыб интенсивность обмена веществ такая же, как и у рыб, обитающих в тропических водах.
Нельзя использовать и один молекулярно-биологический критерий, так как многие макромолекулы (белки и ДНК) обладают не только видовой, но и индивидуальной специфичностью. Поэтому по биохимическим показателям не всегда можно определить, к одному или разным видам относятся особи.
Генетический критерий также не универсален. Во-первых, у разных видов число и даже форма хромосом могут быть одинаковыми. Во-вторых, в одном виде могут быть особи с разным числом хромосом. Так, у одного вида долгоносика имеются диплоидные (2п ), триплоидные (Зп ), тетраплоидные (4п ) формы. В-третьих, иногда особи разных видов могут скрещиваться и давать плодовитое потомство. Известны гибриды волка и собаки, яка и крупного рогатого скота, соболя и куницы. В царстве растений межвидовые гибриды встречаются довольно часто, а иногда бывают и более отдаленные межродовые гибриды.
Нельзя считать универсальным и географический критерий, так как ареалы многих видов в природе совпадают (например,ареал даурской лиственницы и душистого тополя). Кроме того, существуют виды-космополиты, которые распространены повсеместно и не имеют четко ограниченного ареала (некоторые виды сорных растений, комаров, мышей). Ареалы некоторых быстро расселяющихся видов, таких, как домовая муха, изменяются. У многих перелетных птиц различаются ареалы гнездовий и зимовки. Экологический критерий не является универсальным, так как в пределах одного ареала многие виды обитают в очень разных природных условиях. Так, многие растения (например, пырей ползучий, одуванчик) могут жить и в лесу, и на пойменных лугах.
Виды реально существуют в природе. Они относительно постоянны. Виды можно различить по морфологическому, молекулярно-биологическому, генетическому, экологическому, географическому, физиологическому критериям. При определении принадлежности особи к тому или иному виду следует учитывать не один критерий, а весь их комплекс.
Вам известно, что вид состоит из популяций. Популяция представляет собой группу морфологически сходных особей одного вида, свободно скрещивающихся между собой и занимающих определенное место обитания в ареале вида.
Для каждой популяции характерен свой генофонд — совокупность генотипов всех особей популяции. Генофонды разных популяций даже одного вида могут различаться.
Процесс образования новых видов начинается внутри популяции, то есть популяция является элементарной единицей эволюции. Почему же именно популяцию, а не вид или отдельную особь рассматривают как элементарную единицу эволюции?
Особь не может эволюционировать. Она может изменяться, приспосабливаясь к условиям внешней среды. Но эти изменения не эволюционные, так как они не передаются по наследству. Вид, как правило, неоднороден и состоит из ряда популяций. Популяция относительно самостоятельна и может длительное время существовать вне связи с другими популяциями вида. В популяции протекают все эволюционные процессы: у особей возникают мутации, между особями происходит скрещивание, действуют борьба за существование и естественный отбор. В результате генофонд популяции со временем изменяется, и она становится родоначальником нового вида. Именно поэтому элементарная единица эволюции — популяция, а не вид.
Рассмотрим закономерности на следования признаков в популяциях разных типов. Эти закономерности различны для самооплодотворяющихся и раздельнополых организмов. Самооплодотворение особенно часто наблюдается у растений. У самоопыляющихся растений, например гороха, пшеницы, ячменя, овса, популяции состоят из так называемых гомозиготных линий. Чем объясняется их гомозиготность? Дело в том, что при самоопылении увеличивается доля гомозигот в популяции, а доля гетерозигот сокращается.
Чистая линия — это потомки одной особи. Она представляет собой совокупность самоопыляющихся растений.
Начало изучения генетики популяций было положено в 1903 г датским ученым В. Иоганнсеном. Он исследовал популяцию самоопыляемого растения фасоли, легко дающей чистую линию — группу потомков отдельной особи, генотипы которых идентичны.
Иоганнсен взял семена одного сорта фасоли и определил изменчивость одного признака — массы семени. Оказалось, что она варьирует от 150 мг до 750 мг. Ученый высеял отдельно две группы семян: массой от 250 до 350 мг и массой от 550 до 650 мг. Средняя масса семени вновь выросших растений составила в легкой группе 443,4 мг, в тяжелой — 518 мг. Иоганнсен сделал вывод, что исходный сорт фасоли состоит из генетически различных растений.
В течение 6-7 поколений ученый вел отбор семян тяжелых и легких с каждого растения, то есть про водил отбор в чистых линиях. В результате он пришел к выводу, что отбор в чистых линиях не дал сдвига ни в сторону легких, ни в сторону тяжелых семян, значит в чистых линиях отбор не эффективен. А изменчивость массы семян внутри чистой линии является модификационной, ненаследственной и возникает под воздействием условий среды.
Закономерности наследования признаков в популяциях раздельно полых животных и перекрестноопыляемых растений были установлены независимо друг от друга английским математиком Дж Харди и немецким врачом В. Вайнбергом в 1908-1909 гг. Эта закономерность, получившая название закона Харди — Вайнберга, отражает зависимость между частотами аллелей и генотипов в популяциях. Данный за кон объясняет, каким образом в популяции сохраняется генетическое равновесие, то есть число особей с доминантными и рецессивными при знаками остается на определенном уровне.
Согласно этому закону, частоты доминантных и рецессивных аллелей в популяции будут оставаться постоянными из поколения в поколение при наличии определенных условий: высокой численности особей в популяции; свободном их скрещивании; отсутствии отбора и миграции особей; одинаковой численности особей с разными генотипами.
Нарушение хотя бы одного из этих условий ведет к вытеснению одного аллеля (например, А) другим (а). Под действием естественного отбора, популяционных волн и других факторов эволюции особи с доминантным аллелем А будут вытеснять особи с рецессивным аллелем а.
В популяции может измениться соотношение особей с разными генотипами. Предположим, что генетический состав популяции был таким: 20% АА, 50% Аа, 30% аа. Под воздействием факторов эволюции он может оказаться следующим: 40% АА, 50% Аа, 10% аа. Используя закон Харди — Вайнберга, можно вычислить частоту встречаемости любого доминантного и рецессивного гена в популяции, а также любого генотипа.
Популяция — элементарная единица эволюции, так как она обладает относительной самостоятельностью и ее генофонд может изменяться. Закономерности наследования различны в популяциях разных типов. В популяциях самоопыляющихся растений отбор происходит между чистыми линиями. В популяциях раздельнополых животных и перекрестноопыляемых растений закономерности наследования подчиняются закону Харди — Вайнберга.
В соответствии с законом Харди — Вайнберга при относительно постоянных условиях частота аллелей в популяции остается неизменной из поколения в поколение. В этих условиях популяция находится в состоянии генетического равновесия, в ней не происходят эволюционные изменения. Однако в природе нет идеальных условий. Под влиянием факторов эволюции — мутационного процесса, изоляции, естественного отбора и др. — генетическое равновесие в популяции постоянно нарушается, происходит элементарное эволюционное явление — изменение генофонда популяции. Рассмотрим действие различных факторов эволюции.
Один из главных факторов эволюции — мутационный процесс. Мутации были открыты в начале XX в. голландским ботаником и генетиком Де Фризом (1848-1935).
Главной причиной эволюции он считал именно мутации. В то время были известны только крупные мутации, затрагивающие фенотип. Поэтому Де Фриз полагал, что виды возникают в результате крупных мутаций сразу, скачкообразно, без естественного отбора.
Дальнейшие исследования показали, что многие крупные мутации вредны. Поэтому многие ученые считали, что мутации не могут служить материалом для эволюции.
Лишь в 20-х гг. нашего столетия отечественные ученые С. С. Четвериков (1880- 1956) и И. И. Шмальгаузен (1884-1963) показали роль мутаций в эволюции. Было установлено, что любая природная популяция насыщена, как губка, разно образными мутациями. Чаще всего мутации рецессивны, находятся в гетерозиготном состоянии и не проявляются фенотипически. Именно эти мутации и служат генетической ос новой эволюции. При скрещивании гетерозиготных особей эти мутации у потомков могут переходить в гомозиготное состояние. Отбор из поколения в поколение сохраняет особей с полезными мутациями. Полезные мутации сохраняются естественным отбором, вредные — накапливаются в популяции в скрытом виде, создавая резерв изменчивости. Это приводит к изменению генофонда популяции.
Накоплению наследственных различий между популяциями способствует изоляция, благодаря которой между особями разных популяций не происходит скрещивания, а значит, и обмена генетической ин формацией.
В каждой популяции благодаря естественному отбору накапливаются определенные полезные мутации. Через несколько поколений изолированные популяции, обитающие в разных условиях, будут различаться по ряду признаков.
Широко распространена пространственная, или географическая изоляция, когда популяции разделены различными преградами: реками, горами, степями и т. п. Например, даже в близкорасположенных реках обитают разные популяции рыб одного и того же вида.
Различают также экологическую изоляцию, когда особи разных популяций одного вида предпочитают разные места и условия обитания. Так, в Молдавии у желтогорлой лесной мыши образовались лесные и степные популяции. Особи лесных популяций более крупные, пи таются семенами древесных пород, а особи степных популяций — семенами злаков.
Физиологическая изоляция возникает в том случае, когда у особей разных популяций созревание половых клеток происходит в разные сроки. Особи таких популяций не могут скрещиваться. Например, в озере Севан обитают две популяции форели, нерест которых происходит в разные сроки, поэтому они не скрещиваются между собой.
Существует также поведенческая изоляция. Брачное поведение особей разных видов различается. Это препятствует их скрещиванию. Механическая изоляция связана с различиями в строении органов размножения.
Изменение частот аллелей в популяциях может происходить не только под влиянием естественного отбора, но и независимо от него. Частота аллеля может измениться случайным образом. Например, преждевременная гибель особи — единственной обладательницы какого-либо аллеля приведет к исчезновению этого аллеля в популяции. Это явление получило название дрейфа генов.
Важным источником дрейфа генов являются популяционные волны — периодические значительные изменения численности особей популяции. Численность особей изменяется из года в год и зависит от многих факторов: количества пищи, погодных условий, численности хищников, массовых заболеваний и др. Роль популяционных волн в эволюции была установлена С. С. Четвериковым, который показал, что изменение численности особей в популяции влияет на эффективность естественного отбора. Так, при резком сокращении численности популяции могут случайно сохраниться особи с определенным генотипом. Например, в популяции могут сохраниться особи с такими генотипами: 75% Аа, 20% АА, 5% аа. Наиболее многочисленные генотипы, в данном случае Аа, будут определять генный состав популяции до следующей «волны».
Дрейф генов обычно снижает генетическую изменчивость в популяции, главным образом в результате утраты редко встречающихся аллелей. Этот механизм эволюционных изменений особенно эффективен в небольших популяциях. Однако только естественный отбор на основе борьбы за существование способствует сохранению особей с определенным генотипом, соответствующим среде обитания.
Элементарное эволюционное явление — изменение генофонда популяции происходит под влиянием элементарных факторов эволюции — мутационного процесса, изоляции, дрейфа генов, естественного отбора. Однако дрейф генов, изоляция и мутационный процесс не определяют направленности процесса эволюции, то есть выживания особей с определенным, соответствующим среде обитания генотипом. Единственным направляющим фактором эволюции является естественный отбор.
Основные положения эволюционного учения Ч. Дарвина.
Источник